|
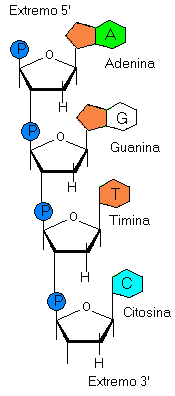
Estructura Primaria del ADN La estructura primaria del ADN es la secuencia de nucleótidos de una sola cadena o hebra, que puede presentarse como un simple filamento extendido o bien algo doblado en sí mismo. Se pueden distinguir en él un esqueleto de fosfopolidesoxirribosa y una secuencia de bases nitrogenadas. El número de hebras de ADN que se puede formar combinando las cuatro bases nitrogenadas (adenina,guanina,citosina y timina) es enorme. Por ejemplo, si el ADN humano tiene 5,6 · 109 pares de nucleótidos, pueden existir 45.600.000.000 ADN diferentes, aunque, en la práctica, existen muchísimos menos. Si se consideran estas innumerables combinaciones posibles, se puede comprender cómo a través de la secuencia de bases nitrogenadas es posible estructurar una determinada información, el llamado mensaje biológico o información genética. De la misma manera, con veintiseis letras se estructura la información intelectual o lenguaje. Con estas letras, tomadas unas veces individualmente y otras en grupas de dos, tres, cuatro, etc., y combinándolas conveninetemente, se ha escrito, por ejemplo, viente poemas de amor y una canción desesperada |
 |
 |
Estructura Secundaria La estructura secundaria del ADN es la disposición en el espacio en dos hebras o cadenas de polinucleótidos en doble hélice, con las bases nitrogendas enfrentadas y unidas mediante puentes de hidrógeno. |
|
a) La densidad y viscosidad de las dispersiones acuosas del ADN eran distintas de las esperadas, es decir, de las que se habían calculado a partir de su composición química. Por tanto, los grupos -NH2 , -CO- y =NH deberían establecer puentes de hidrógeno entre si. En la estructura secundaria del ADN, al igual que en las proteínas, los grupos hidrófobos =C-H y -CH3 de las bases se disponen hacia el interior de la molécula, estableciéndose interacciones hidrofóbicas entre grupos lipófilos, que colaboran con los puentes de hidrógeno en dar estabilidad a la macromolécula. Las pentosas y los grupos fosfato quedan en el exterior. Debido a la ionización de estos últimos, los ácidos nucleicos tiene carácter ácido y se definen como polianiones. La doble hélice de ADN en estado natural es muy estable; pero, si se calienta una dispersión de fibras de ADN, cuando la temperatura llega aproximadamente a 100 ºC, las dos hebras de la doble hélice se separan, es decir, se produce la desnaturalización del ADN. Si posteriomente se mantiene el ADN desnaturalizado a 65 ºC, las dos hebras vuelven a unirse. Esta restauración de la doble hélice es lo que se llama renaturalización o hibridación del ADN. En la actualidad se conocen tres tipos de estructura de doble hélice del ADN: las formas B, A y Z
|